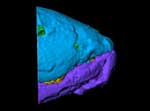
Eusthenopteron foordi is one of the most scientifically important fossil vertebrates. It is a fossil lobe-finned (Sarcopterygian) fish, which belongs in the stem group of the Tetrapoda or land vertebrates. This means that it is more closely related to land vertebrates to us than any living fish. The two still surviving lobe-finned fish groups, the lungfishes or Dipnoi (see Chirodipterus) and the coelacanths of Actinistia (see Latimeria), are both more distantly related to us. Eusthenopteron lived during the earlier part of the Late Devonian period, about 370 million years ago.
Eusthenopterons fame rests partly on its phylogenetic position as a close tetrapod relative, and partly on the fact that it was the chosen subject of one of the most painstaking investigations of a fossil vertebrate ever undertaken. Over the course of nearly four decades, ending with a monographic description in 1980, Professor Erik Jarvik of the Swedish Museum of Natural History (Naturhistoriska Riksmuseet) in Stockholm, Sweden, dissected the complete anatomy of a perfectly preserved specimen of Eusthenopteron from Miguasha in Quebec, Canada. To study the head he used the serial grinding technique, whereby the specimen was ground down to a flat surface, a fraction of a millimeter at a time, and each ground surface was drawn (greatly enlarged) and photographed before being ground away to reveal the next, deeper, section through the skull. By cutting out thin sheets of wax to match the bone outlines in the drawings, and layering these on top of each other, Jarvik was able to build complete wax models, not only of the bones of the head, but also of the internal spaces that enclosed the brain and sense organs; results similar to those that we can now produce with CT scanning. However, Jarviks grinding work took more than a decade to complete, as against a few hours for a CT scan, and the specimen was destroyed in the process.
Our specimen of Eusthenopteron also comes from Miguasha and represents one of the largest individuals recovered from that site. Fossils of Eusthenopteron are also known from other parts of the world, such as Russia and the Baltic States, but these are generally much less complete. Our fossil comprises the skull, gill region, shoulder girdle, and one pectoral fin; some of the external bones have flaked off and the skull has become slightly asymmetrical because of compression in the sediment, but in other respects it is very well preserved. The internal bones of the head, such as the palate and braincase, are less dense than the external bones and show up only faintly on the cutaways and skeleton movies. However, they can be clearly seen on the slice movies, and a separate rendering project for these elements is under way (see 'Additional Imagery').
Three key features of Eusthenopteron can be seen in these images:
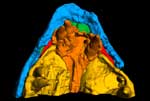
 The internal nostril or choana, which opens from the nasal sac onto the palate, can be seen clearly on the right side in coronal slice images 32-40. This is one of the characters that unites Eusthenopteron with tetrapods: in most other fishes, the nasal sac has two external nostrils but no connection with the mouth.
The internal nostril or choana, which opens from the nasal sac onto the palate, can be seen clearly on the right side in coronal slice images 32-40. This is one of the characters that unites Eusthenopteron with tetrapods: in most other fishes, the nasal sac has two external nostrils but no connection with the mouth.
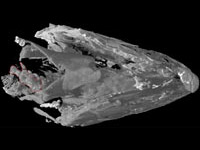 The pectoral fin skeleton on the left side of the fish is visible in the rotating movies and in coronal slice images 269-334 (where it lies on the right). The internal skeleton of the fin has a structure similar to our arm, with a single proximal element (the humerus) supporting two parallel elements (radius and ulna) and a number of smaller bones at the distal end corresponding to our wrist bones. There is no equivalent of the hand: the numerous thin, bright, more or less parallel rods at the distal margin of the skeleton are "lepidotrichia" or dermal fin rays, comparable to those that form the bulk of the fin in an actinopterygian (ray-finned bony fish) such as Halichoeres. In ray-finned fishes the endoskeleton is small and does not resemble a limb. Lungfishes and coelacanths (the two living lobe-fin groups) do have limb-like fin skeletons, but the match to tetrapod limb structure is less precise than in Eusthenopteron.
The pectoral fin skeleton on the left side of the fish is visible in the rotating movies and in coronal slice images 269-334 (where it lies on the right). The internal skeleton of the fin has a structure similar to our arm, with a single proximal element (the humerus) supporting two parallel elements (radius and ulna) and a number of smaller bones at the distal end corresponding to our wrist bones. There is no equivalent of the hand: the numerous thin, bright, more or less parallel rods at the distal margin of the skeleton are "lepidotrichia" or dermal fin rays, comparable to those that form the bulk of the fin in an actinopterygian (ray-finned bony fish) such as Halichoeres. In ray-finned fishes the endoskeleton is small and does not resemble a limb. Lungfishes and coelacanths (the two living lobe-fin groups) do have limb-like fin skeletons, but the match to tetrapod limb structure is less precise than in Eusthenopteron.
thumb.jpg) The intracranial joint is a peculiar feature of lobe-finned fishes that has been lost in tetrapods. It is still present in the coelacanth Latimeria, but lungfishes lost it very early in their evolution, as shown by its absence in Chirodipterus. The external part of the joint, between the parietal and postparietal bones, can be seen as a dark transverse line across the middle of the skull roof in the pitch and roll movies. The internal, braincase component is best seen in the horizontal slice movie. The joint probably served to allow vertical movement of the snout during feeding.
The intracranial joint is a peculiar feature of lobe-finned fishes that has been lost in tetrapods. It is still present in the coelacanth Latimeria, but lungfishes lost it very early in their evolution, as shown by its absence in Chirodipterus. The external part of the joint, between the parietal and postparietal bones, can be seen as a dark transverse line across the middle of the skull roof in the pitch and roll movies. The internal, braincase component is best seen in the horizontal slice movie. The joint probably served to allow vertical movement of the snout during feeding.

The specimen was scanned by Matthew Colbert and Richard Ketcham on 16 February 2004 along the coronal axis for a total of 340 slices. Each 1024x1024 pixel slice is 0.8 mm thick, with an interslice spacing of 0.8 mm and a field of reconstruction of 169 mm.
A second partial scan, extending from the tip of the snout to the orbit, was performed with an interslice spacing and slice thickness of 0.4 mm and a field of reconstruction of 169 mm. This close-up scan, consisting of 250 1024x1024 pixel slices, was used to produce the first three animations on the 'Additional Imagery' page.



 ,
,  )