This page serves supplemental imagery for a paper entitled Assembling the Squamate Tree of Life: Perspectives from the Phenotype and the Fossil Record by J.A. Gauthier, M. Kearney, J.A. Maisano, O. Rieppel, and A.D.B. Behlke (Bulletin of the Peabody Museum of Natural History, 53, 3-308, 2012). The abstract is as follows:
We assembled a dataset of 192 carefully selected species 51 extinct and 141 extant and 976 apomorphies distributed among 610 phenotypic characters to investigate the phylogeny of Squamata (lizards, including snakes and amphisbaenians). These data enabled us to infer a tree much like those derived from previous morphological analyses, but with better support for some key clades. There are also several novel elements, some of which pose striking departures from traditional ideas about lizard evolution (e.g., that mosasaurs and polyglyphanodontians are on the scleroglossan stem, rather than parts of the crown, and related to varanoids and teiids, respectively). Long-bodied, limb-reduced, snake-like fossorial lizards most notably dibamids, amphisbaenians and snakes have been and continue to be the chief source of character conflict in squamate morphological phylogenetics. Carnivorous lizards (especially snakes, mosasaurs and varanoids) have proven a close second. Genetic data, presumably less burdened by the potential for adaptive convergence related to fossoriality, were expected to resolve these conflicts. Although recent gene phylogenies seem to do so, they also differ radically from any phylogeny based on the phenotype, especially for the most ancient crown squamate divergences that occurred during the latter half of the Mesozoic. Our study relied on traditionally prepared specimens as well as high-resolution computed tomography scans that afforded unprecedented access to the cranial anatomy of Squamata. This, along with the inclusion of stem fossils, provided an unparalleled sample of the phenotype enabling us to more fully explore the extreme incongruences between molecular and morphological topologies for the squamate tree of life. Despite this extensive new database, we were unable to find morphological support for the major rearrangement of the deep divergences in Squamata proposed by recent molecular studies. Instead, our data strongly support the same fundamental topology suggested by most previous morphological studies an Iguania-Scleroglossa basal split, a sister-group relationship between Gekkota and Autarchoglossa, and the divergence between Anguimorpha and Scincomorpha and documents the extreme degree of morphological homoplasy required by those molecular topologies.

About the Species
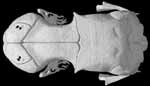
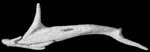
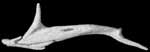
About this Specimen
The specimen was scanned by Matthew Colbert on 11 February 2005 along the coronal axis for a total of 570 slices. Each 1024x1024 pixel slice is 0.016 mm thick, with an interslice spacing of 0.016 mm and a field of reconstruction of 5.5 mm.
About the
Scan
Literature
Evans, H. E. 1955. The osteology of a worm snake, Typhlops jamaicensis (Shaw). Anatomical Record 122:381-396.
Iordansky, N. N. 1997. Jaw apparatus and feeding mechanics of Typhlops (Ophidia: Typhlopidae): a reconsideration. Russian Journal of Herpetology 4:120-127.
Kley, N. J. 2001. Prey transport mechanisms in blindsnakes and the evolution of unilateral feeding systems in snakes. American Zoologist 41:1321-1337.
List, J. C. 1966. Comparative osteology of the snake families Typhlopidae and Leptotyphlopidae. Ill. Biological Monographs 36:1-112.
McDowell, S. B. 1967. Osteology of the Typhlopidae and Leptotyphlopidae: a critical review. Copeia 1967:686-692.
Rieppel, O. 1979. The braincase of Typhlops and Leptotyphlops (Reptilia: Serpentes). Zoological Journal of the Linnean Society 65:161-176.
Links
Typhlops jamaicensis page on The Reptile Database
Literature
& Links
Three-dimensional volumetric renderings of the skull with the jaw removed and of the isolated left mandible. All are less than 2mb.
Additional
Imagery




 )