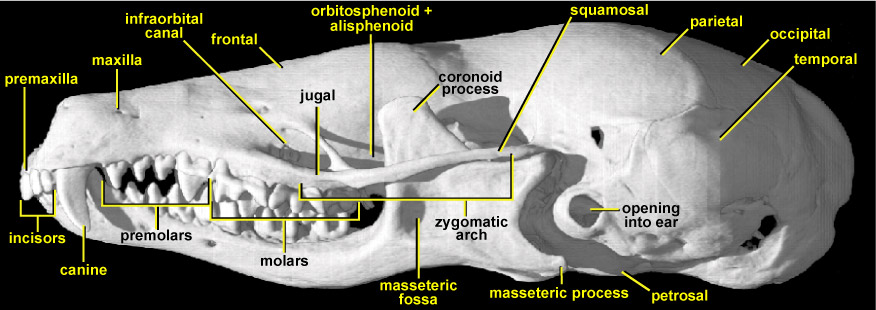
This view of the European mole skull shows several functionally significant features. The rostrum (muzzle) is very long and rather low. Its sides and lower tooth-bearing border are composed of the premaxilla at the very front and the long maxilla behind; this lower border is upwardly concave. Towards the posterior end of the rostrum, on its lateral face and just in front of the orbit, there is a triangular opening. This is the infraorbital canal. It is very large in Talpa because it gives admission to the massive infraorbital nerve that supplies the sensory fields on its nose, such as tactile vibrissae (hairs) and the soft sensitive skin of the nose itself. The infraorbital canal passes through the maxilla and has the jugal underneath it. The zygomatic arch (cheekbone) is extremely thin, being reduced to a mere splint; its front half is made up of the jugal whereas its rear half is made up by the squamosal. This reduction suggests a similar reduction in the size of the masseter (chewing) muscle that attaches to it and then connects to the lower rear part of the lower jaw. However, the large masseteric process (the backward-pointing splint at the rear lower corner of the lower jaw onto which the masseter attaches) and the sizeable masseteric fossa shows that the masseter muscle in T. europaea is large. Indeed this powerful jaw-closing muscle originates mainly from the jugal, beneath the orbit on the side of the snout, just underneath the infraorbital canal. The large size of the masseteric process is also biomechanically important because when the jaws open wide, the process swings backwards and upwards and abuts against the petrosal. This prevents the jaws from being opened very wide.
In this view, the high pointed crowns of the cheek teeth can be seen, as well as the curved blade-like canine that lies at the front of the jaws. The lower jaw has a tall, triangular, plate-like blade that passes upwards between the side of the main part of the skull and the zygomatic arch. This triangular plate is the coronoid process of the lower jaw. In Talpa it is very substantial, indicating the attachment area needed for a correspondingly large temporalis muscle. The temporalis muscle is the main jaw-closing muscle in Talpa; it also attaches to the surface of the rounded part of the skull that houses the brain, towards the rear of the skull. The coronoid process of Talpa is highly unusual for a mammal in that it is inclined anteriorly, rather than vertically, towards the orbital region. This increases its mechanical advantage; producing more force at the teeth per unit force of temporalis contraction (Crompton, 1963). However this geometry would potentially cause over-stretching of the temporalis fibres. In life this does not happen because the jaws do not open very wide owing to the arrangement of the masseteric process.
The orbit is roofed by the frontal bone, while the orbitosphenoid and alisphenoid bones lie below the frontal on the side of the skull. The latter two bones are to a large extent hidden in lateral view by the tall coronoid process, and because they often fuse, they are often impossible to make out as individual bones. There is a prominent bulge of the braincase at the orbital region (just by the tip of the coronoid process, better seen in dorsal view). In most mammals this region is constricted in order to make room for the eyeballs. Because eyesight in Talpa is severely reduced, its eyeballs are tiny, so there is no need for the braincase to narrow here. The braincase in Talpa is greatly expanded, particularly bilaterally; this gives a flattened but bulbous appearance. Most of this expanded region at the rear of the skull is made up by the parietals, which form its roof, with the occipital at the rear. The temporalis muscle originates from this broad surface and runs forward horizontally to insert on the coronoid process. The temporalis muscle therefore has a high mechanical leverage and gives Talpa an unusually powerful bite for such a small mammal. This strong bite relates to the increasing material toughness of the chitinous exoskeleton of its invertebrate prey as a scaling function of its small size. A powerful bite is therefore required to puncture-crush the exoskeleton of beetles that may be up to 20% of its body size: biomechanically this is like a lion having to bite through a bug the size of a sheep! Additionally the strong bite may relate to underground territorial combats between rivals during the mating season in an environment where escape is difficult and the ability to discourage a combatant is at a premium. In lateral view, the cusps of the molar and premolar teeth in Talpa are very tall and sharp.
Because Talpa digs using its forelimbs in loose soil, the skull is not adapted directly as a tunnelling tool. Its cranial anatomy contrasts with that of the Australian marsupial mole, Notoryctes, which swims through the subsurface of loose sand using its head as a shovel. Its skull is high and wedge-shaped, whereas that of Talpa is low and broad, as is its brain. Despite the long cavum nasi (nasal cavity) of Talpa, its olfactory system has a lower cell density and fewer receptor cells than many of its insectivore relatives. But in searching for food, the use of the nose as a tactile organ is extremely important; almost above all the moles other senses. Eyesight is very strongly reduced in Talpa, but it has better visual acuity than most other moles. The large opening into the ear of the mole can be seen just behind the jaw joint at the side of the skull. This hole passes through the temporal bone. Although moles have relatively poor smell, their nasal turbinate (scroll) bones, which are associated with smell, occupy not just the whole of the nasal cavity but extend back into the rear portions of the skull as well, as far as the posterior part of the orbital region. These convoluted internal bones can be seen in the coronal CT slices of Talpas skull.