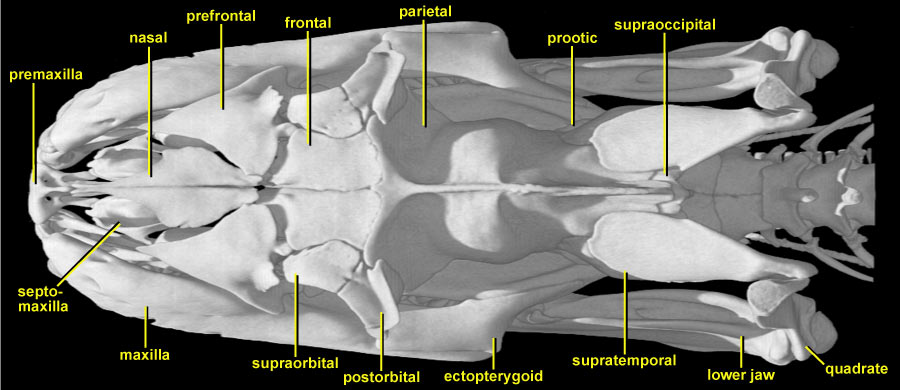
In dorsal view, the Python skull is seen to be long and rather narrow. Running from rostral to caudal along the midline are: the small single premaxilla; rhomboid nasals; short frontals; and a long crested median parietal. The tip of the supraoccipital is just visible behind the end of the parietal, and then the skull connects with the vertebral column. Bilaterally, the borders of the skull are provided by the maxillae and the ectopterygoids. The rear of the lower jaw is then visible on each side extending posteriorly to its articulation with the quadrate.
Beginning anteriorly, note the pair of small bones (mostly in shadow) that lie behind the premaxilla and under the nasals, curving upwards slightly on each side. These are the septomaxillae that enclose and support the vomeronasal apparatus (accessory taste/olfactory system the forked tongue tips transmit chemical particles into these organs in the roof of the mouth, thus the snake seems to be tasting the air). The nasals taper anteriorly and posteriorly. Note the weak contact between the nasal and frontal. This is one of the kinetic joints of the skull allowing bending within the skull table (prokinesis). Coming into the angle between the nasals and frontals on each side is a larger triradiate prefrontal that forms the anterior margin of the small orbit. The frontal itself is excluded from the orbital margin by the abutment of the prefrontal with a small bone labelled postfrontal by Parker and Grandison (1977), but supraorbital by Frazzetta (1959). Rieppel and Zaher (2000) state that the postfrontal is absent in snakes (their character 23), and therefore supraorbital is used here. Posterolaterally, the supraorbital contacts the larger postorbital, forming the rear margin of the orbit. The massive parietal provides a major component of the skull roof, extending both laterally and ventrally to enclose and protect the brain (see also slices). Posterolaterally, it is overlapped by the supratemporals. The contact permits movement between the supratemporal and the skull roof so that the posterior ends of the supratemporals can swing out laterally to increase the width of the skull. At its tip, each supratemporal meets (and suspends) the quadrate in a highly mobile joint (streptostyly). This combination of joints (mobile supratemporal/parietal/prootic, supratemporal/quadrate, quadrate/lower jaw) permits the extended gape that typifies higher snakes (macrostomatan big-mouthed). In front of the anterior rim of the supratemporal, and below the parietal, the tip of the prootic is just visible on each side. There is no squamosal.
Perhaps the most important thing to grasp from this view is the sharp division of the skull into two functional parts: those parts concerned with feeding and those parts protecting the brain and major sense organs. Contrast, for example, the rather delicately built anterior part where the sutures are open, and the very strong frontoparietal complex which supports and protects the brain. This allows the snake to have a very dynamic skull in terms of flexibility while feeding, while at the same time ensuring that the brain and major sense organs are not damaged. Thus the feeding apparatus is suspended (both anteriorly and posteriorly) from the central strongbox of the compound braincase (see also slices).