Diomedea immutabilis, although still a large bird, is small in size for an albatross (only 79-81 cm long, compared to other albatrosses that may reach sizes of 125 cm in length). When observed in the air, albatrosses appear incredibly gracefully, using a combination of 'dynamic' and 'slope' soaring; however, their landing often may be described as somewhat clumsy. As with other albatrosses D. immutabilis is strictly a marine bird, for the most part only going to land to breed. They feed primarily on squid and other small marine animals and even sleep while floating on the water. These large birds may live up to 40 years and do not even begin to breed until they reach an age of eight to nine years. Because they spend most of their lives at sea, little is known about the natural history of albatrosses compared with other birds. However, the breeding behavior of the Laysan albatross is well documented and includes an elaborate courtship 'dance' (Whittow, 1993). |
|
The biogeographic range of the Laysan albatross extends throughout the northern part of the Pacific ocean during most of the year. Breeding colonies are established on parts of the Hawaiian archipelago as well as islands off of the coasts of Mexico and Japan (Whittow, 1993).
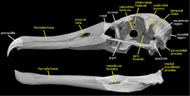
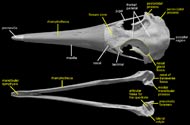
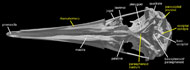
The albatrosses are traditionally included with the Procellariiformes, which also includes shearwaters, petrels, and fulmars (Wetmore, 1960). This classification of Procellariiformes was considered to be monophyletic by Cracraft (1988) and placed as the sister taxon to 'Pelecaniformes' (a somewhat more questionable taxon). These two groups were joined by a number of cranial characters (observable in the labeled images of the skull), including a greatly enlarged upper tympanic recess, bony nostrils that have been reduced to small openings, a rostrum with long nasal grooves, a terminal hook on the bill, and a large, ventrally exposed pneumatic fenestra that lies between the fenestra ovalis and the articular surface for the medial condyle of the quadrate (Cracraft, 1988). The position of procellariiforms within Neoaves, however, has not been stable. More recent DNA-DNA hybridization studies show a sister-group relationship between traditional procellariiforms and sphenisciforms (penguins) (Van Tuinen, 2001); whereas morphological studies place them as most closely related to grebes and flamingoes (Mayr and Clarke, 2003).
The most distinctive feature of the albatross skull is the fossa that houses the nasal gland. This gland (sometimes referred to as the salt gland) is used to maintain electrolyte homeostasis and water balance during conditions of salt-loading (Siegel-Causey, 1990). Although it is typically more pronounced in marine species that intake a large amount of salt, such as albatrosses, many terrestrial birds also possess a nasal gland. The feature is recognized in osteological specimens by a marked depression on the anterior-most portion of the frontal bone. This character has been used to join the grebes, loons, penguins and albatrosses (Mayr and Clarke, 2003); however, in pelecaniforms it is present within the orbit itself on the orbital surface of the frontal (Siegel-Causey, 1990).
Labeled 3D Renderings:

About the Species
This specimen (FMNH 313780) was made available to the University of Texas High-Resolution X-ray CT Facility for scanning by Dr. Timothy Rowe of the Department of Geological Sciences, The University of Texas at Austin. Funding for image processing was provided by a National Science Foundation Digital Libraries Initiative grant to Dr. Rowe.
About this Specimen
The specimen was scanned by Matthew Colbert on 28 September 2004 along the coronal axis for a total of 1275 slices, each slice 0.138 mm thick with an interslice spacing of 0.138 mm.
About the
Scan
Literature
Baumel, J. J., A. S. King, J. E. Breazile, H. E. Evans, and J. C. Vanden Berge (eds.). 1993. Handbook of Avian Anatomy: Nomina Anatomica Avium, Second Edition. Publication of the Nuttall Ornithological Club, number 23. Nuttall Ornithological Club, Cambridge, Massachusetts, 779 pp.
Cracraft, J. 1988. The major clades of bird; pp. 333-355 in M. J. Benton (ed.), The Phylogeny and Classification of the Tetrapods. Clarendon Press, Oxford.
Mayr, G., and J. Clarke. 2003. The deep divergences of neornithine birds: a phylogenetic analysis of morphological characters. Cladistics 19:527-553.
Siegel-Causey, D. 1990. Phylogenetic patterns of size and shape of the nasal gland depression in Phalacrocoracidae. Auk 108:110-118.
Van Tuinen, M., D. B. Butvill, J. A. W. Kirsch, and S. B. Hedges. 2001. Convergenece and divergence in the evolution of aquatic birds. Proceedings of the Royal Society, London B, 268:1345-1350.
Wetmore, A. 1960. A classification for the birds of the world. Smithsonian Miscellaneous Collections 139:1-37.
Whittow, G. C. 1993. Laysan Albatross. Birds of North America 66:1-20.
Links
The Laysan albatross on the USGS Patuxent Wildlife Research Center website
A species profile of Diomedea immutabilis on project OBIS-SEAMAP. (Ocean Biogeographic Information System - Spatial Ecological Analysis of Megavertebrate Populations)
Photos of Diomedea immutabilis on the Hawaiian Ecosystems at Risk Project.
Literature
& Links
None available.
Additional
Imagery
|



 )