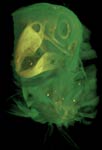
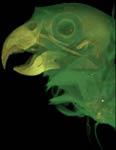
Brotogeris chrysopterus, the golden-winged parakeet, is a member of Psittaciformes (parrots), a group that ranges throughout tropical and into temperate climates in both the Old and New World. The geographic range of B. chrysopterus, however, is restricted primarily to eastern Brazil.
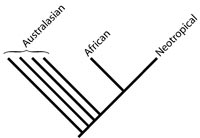
Little controversy surrounds the monophyly of extant Psittaciformes. The group is united by a number of diagnostic characters, including the presence of two unique muscles (M. ethmomandibularis and M. pseudomasseter) along with a suborbital arch (Tokita, 2004; Burton, 1974; Beecher, 1962). The position of parrots within crown-group Aves is somewhat more contentious. Recent morphological and total evidence studies place Psittaciformes as most closely related to Coliiformes (mousebirds; Mayr and Clarke, 2003; Mayr et al., 2002). Other analyses place them as the sister taxon to members of 'Coraciiformes' (morphological; Livezey and Zusi, 2001) or the sister taxon to a large clade comprised of an assemblage of neognath birds (DNA-DNA hybridization; Sibley and Ahlquist, 1990). For an overview of the classification and taxonomy of Psittaciformes see Livezey and Zusi (2001). In general, parrots are characterized by a hooked bill, zygodactylous feet, unique intestinal looping, lack of caeca, presence of an aftershaft on feathers, and a diastatic wing (Smith, 1975). Although the group at one time was broken into two major groups: Cacatuoidea (Australasian, Oriental, Neotropical, and Ethiopian) and Platycercoidea (Australasian and Neotropical) (Smith, 1963), more recent phylogenetic analyses of molecular data divide parrots up into groups that are more consistent with present day geography (Miyaki et al., 1998). The Neotropical and African species are each monophyletic and represent each others closest relatives. Australasian parrots are a paraphyletic assemblage with respect to the aforementioned taxa (Miyaki et al, 1998).
Much of the unique cranial morphology of parrots is a product of the increased kinesis of the skull. Although still categorized as prokinetic, the skull possesses many additional modifications to help manipulate and shell food, including the presence of several new muscles. The pseudomasseter, unique to parrots, is a branch of the anterior adductor mandibulae externus ventralis. This offshoot attaches to the suborbital bar, another novel feature (osteological) that is present within several lineages of parrots (Zusi, 1993). Parrots also have a unique ethmomandibularis muscle that attaches to the interorbital septum (Tokito, 2004; Burton, 1974). These muscles are innervated by the mandibular branch of the trigemeninal nerve (Tokita, 2004; Burton, 1974; Beecher, 1962). The morphology of the hyoid apparatus is also highly specialized to support the unusually agile tongue, which aids in the most famous characteristic of parrots, speech.
The fossil record of parrots stretches back at least as far as the Eocene of Europe. Stem group psittaciforms are known from material that comes from the Early Eocene London Clay Formation, England (Dyke and Cooper, 2000), as well as articulated material from the Middle Eocene formation at Messel, Germany (Mayr, 2002). An isolated record of a lower jaw of a psittaciform was reported from the Cretaceous Lance Formation, Wyoming (Stidham, 1998). This edentulous jaw fragment is allocated to Psittaciformes based primarily on the k-shaped pattern formed by the neurovascular canals and the deeply concave symphysis (Stidham, 1998). This find has larger significance for Neornithes as a whole. If the fragment belongs to a parrot, it would provide further evidence that Neornithes diverged in the Cretaceous rather than the Eocene (Clarke et al., 2005).
Labeled Renderings:

Literature
Baumel, J. J., A. S. King, J. E. Breazile, H. E. Evans, and J. C. Vanden Berge (eds.). 1993. Handbook of Avian Anatomy: Nomina Anatomica Avium, Second Edition. Publication of the Nuttall Ornithological Club, number 23. Nuttall Ornithological Club, Cambridge, Massachusetts, 779 pp.
Beecher, W. J. 1962. The bio-mechanics of the bird skull. Bulletin of the Chicago Academy of Sciences 11:10-33.
Burton, P. J. K. 1974. Jaw and tongue features in Psittaciformes and other orders with special reference to the anatomy of the tooth-billed pigeon (Didunculus strigirostris). Journal of Zoology 174:255-276.
Clarke, J. A., C. P. Tambussi, J. I. Noriega, G. M. Erickson, and R. A. Ketcham. 2005. Definitive fossil evidence for the extant avian radiation in the Cretaceous. Nature 433:305-308.
Dyke, G. J., and J. H. Cooper. 2000. A new psittaciform bird from the London Clay (Lower Eocene) of London. Paleobiology 43:271-285.
Livezey, B. C., and R. L. Zusi. 2001. Higher-order phylogenetics of modern Aves based on comparative anatomy. Netherlands Journal of Zoology 51:179-205.
Mayr, G. 2002. On the osteology and phylogenetic affinities of the Pseudasturidae -- Lower Eocene stem-group represetatives of parrots (Aves, Psittaciformes). Zoological Journal of the Linnean Society 136:715-729.
Mayr, G., and J. Clarke. 2003. The deep divergences of neornithine birds: a phylogenetic analysis of morphological characters. Cladistics 19:527-553.
Mayr, G., A. Manegold, and U. S. Johansson. 2003. Monophyletic groups within 'higher land birds' -- comparison of morphological and molecular data. Journal of Zoological Systematics and Evolutionary Research 41:233-248.
Mikyaki, C. Y., S. R. Matioli, T. Burke, and A. Wajntal. 1998. Parrot evolution and paleogeographical events: mitochondrial DNA evidence. Molecular Biology and Evolution 15:544-551.
Sibley, C. G., and J. E. Ahlquist. 1990. Phylogeny and Classification of Birds: A Study in Molecular Evolution. Yale University Press, New Haven, Connecticut.
Smith, G. A. 1975. Systematics of parrots. Ibis 117:18-68.
Stidham, T. 1998. A lower jaw from a Cretaceous parrot. Nature 396:29-30.
Tokita, M. 2004. Morphogenesis of parrot jaw muscles: understanding the development of an evolutionary novelty. Journal of Morphology 259:69-81.
Zusi, R. 1993. Patterns of diversity in the avian skull; in J. Hanken and B. K. Hall (eds.), The Skull, Volume 2. University of Chicago Press, Chicago, Illinois.
Links
A detailed report of Brotogeris chrysopterus on InfoNatura.



 )



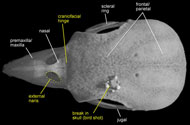
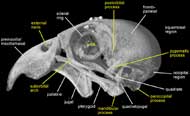
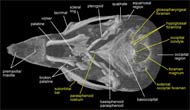

)
