Gastropods (such as snails and slugs) are the most diverse group in the phylum Mollusca. With more than 40,000 living representatives, gastropods also have an extensive fossil record that reaches well into the Cambrian (560 Ma). While their body form is the least modified from the ancestral molluscan body plan (similar to a cap-like limpet form), they occupy the widest variety of habitats and have successfully infiltrated the plankton, intertidal zone, nearshore to abyssal marine, freshwater and terrestrial (modified lung to breathe air on land) realms. A shell may be absent or present and shell form may be cap-like (limpets and abalone), spiraled (cones), and very ornate (some conchs). The radula (a conveyor belt of tiny teeth) allows gastropods to exploit a variety of diets including algae grazing (limpets), herbivory, scavenging, and predatory drilling (neogastropods). In the genus Conus, the radula is so highly modified that this predatory gastropod captures its prey by ejecting the radula like a harpoon.
Perhaps the most peculiar aspect of gastropods is the phenomenon of torsion. During the larval stage, veligers (molluscan larvae with swimming appendages) initially develop so that the anus and gills are at the rear of the body, but later the body contorts so that the anus and gills lie directly over the gastropods head. While a number of hypotheses have been suggested, there appears to be no simple explanation for this complicated developmental feature.
|
Athleta tuomeyi, a neogastropod of the family Volutidae, ranges from the Late Paleocene to Early Eocene of the Atlantic and Gulf Coastal Plains from Virginia to Mexico. What distinguishes this species from the rest of the Athleta petrosa stock (Fisher et al., 1964) is the development of an extreme parietal callus. Curiously, some other molluscs (bivalves and other gastropods) are known to develop extreme callus on the parietal area of the shell. A number of hypotheses have been invoked to explain this unusual accretionary growth, including protection from increased predation, additional stabilizing weight for high-energy nearshore environments, and disease. |

About the Species
These specimens were collected from the Early Eocene, Woods Bluff, Alabama. They were made available to The University of Texas High-Resolution X-ray CT Facility for scanning by Heather Ford of San Diego State University. Funding for image processing was provided by a National Science Foundation Digital Libraries Initiative grant to Dr. Timothy Rowe of The University of Texas at Austin.

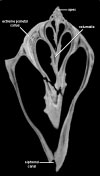
Dorsal view of one of the scanned specimens.
About this Specimen
The specimens were scanned by Matthew Colbert on 2 September 2005 along the coronal axis for a total of 465 1024x1024 pixel slices. Each slice is 0.113 mm thick, with an interslice spacing of 0.113 mm and a field of reconstruction of 54 mm.
About the
Scan
Dockery, D. 1998. Molluscan faunas across the Paleocene/Eocene series boundary in the North American Gulf Coastal Plain, p. 296-322. In Aubry, M.-P., Lucas, S. G., and Berggren, W. (eds.), Late Paleocene-early Eocene climatic and biotic events in the marine and terrestrial records. Columbia University Press, New York.
Fisher, W. L., Rodda, P. U., and Dietrich, J. W. 1964. Evolution of Athleta petrosa stock (Eocene, Gasropoda of Texas). University of Texas, Bureau of Economic Geology.
Prothero, D. R. 2004. Bringing Fossils To Life: An Introduction To Paleobiology: 2nd Edition. McGraw-Hill, Boston, 503 pp.
Literature
& Links
Front page image.
|  |
Additional
Imagery
|



 ,
,  )



)